
la celula
27.10.2014 15:28La célula
Se define a la célula como la unidad morfológica y funcional de todo ser vivo. De hecho, la célula es el elemento de menor tamaño que puede considerarse vivo. Como tal posee una membrana de fosfolípidos con permeabilidad selectiva que mantiene un medio interno altamente ordenado y diferenciado del medio externo en cuanto a su composición, sujeta a control homeostático, la cual consiste en biomoléculas y algunos metales y electrolitos. La estructura se automantiene activamente mediante el metabolismo, asegurándose la coordinación de todos los elementos celulares y su perpetuación por replicación a través de un genoma codificado por ácidos nucleícos. La parte de la biología que se ocupa de ella es la citología.
Las células son los módulos básicos de todos los tejidos y los órganos de los seres humanos y de todos los otros organismos. Aparecieron hace 3500 millones de año.
Las células, como sistemas termodinámicos complejos, poseen una serie de elementos estructurales y funcionales comunes que posibilitan su supervivencia; no obstante, los distintos tipos celulares presentan modificaciones de estas características comunes que permiten su especialización funcional y, por ello, la ganancia de complejidad. De este modo, las células permanecen altamente organizadas a costa de incrementar la entropía del entorno, uno de los requisitos de la vida.
Descubrimiento
A continuación se enumera una breve cronología de tales descubrimientos:
-
1665: Robert Hooke publicó los resultados de sus observaciones sobre tejidos vegetales, como el corcho, realizadas con un microscopio de 50 aumentos construido por él mismo. Este investigador fue el primero que, al ver en esos tejidos unidades que se repetían a modo de celdillas de un panal, las bautizó como elementos de repetición, «células» (del latín cellulae, celdillas). Pero Hooke sólo pudo observar células muertas por lo que no pudo describir las estructuras de su interior.
-
Década de 1670: Anton van Leeuwenhoek, observó diversas células eucariotas (como protozoos y espermatozoides) y procariotas (bacterias).
-
1745: John Needham describió la presencia de «animálculos» o «infusorios»; se trataba de organismos unicelulares. Dibujo de la estructura del corcho observado por Robert Hooke bajo su microscopio y tal como aparece publicado en Micrographia.
-
Década de 1830: Theodor Schwann estudió la célula animal; junto con Matthias Schleiden postularon que las células son las unidades elementales en la formación de las plantas y animales, y que son la base fundamental del proceso vital.
-
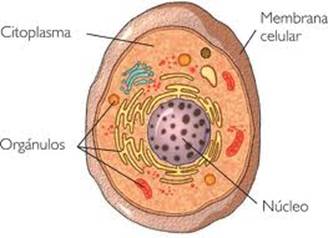
1831: Robert Brown describió el núcleo celular.
-
1839: Purkinje observó el citoplasma celular.
-
1850: Rudolf Virchow postuló que todas las células provienen de otras células.
-
1857: Kölliker identificó las mitocondrias.
-
1860: Pasteur realizó multitud de estudios sobre el metabolismo de levaduras y sobre la asepsia.
-
1880: August Weismann descubrió que las células actuales comparten similitud estructural y molecular con células de tiempos remotos.
-
1931: Ernst Ruska construyó el primer microscopio electrónico de transmisión en la Universidad de Berlín. Cuatro años más tarde, obtuvo un poder de resolución doble a la del microscopio óptico.
-
1981: Lynn Margulis publica su hipótesis sobre la endosimbiosis serial, que explica el origen de la célula eucariota.
Teoría celular
En 1830 se disponía ya de microscopios con una óptica más avanzada, lo que permitió a investigadores Theodor Schwann y Matthias Schleiden definir los postulados de la teoría celular, la cual afirma, entre otras cosas:
-
Que la célula es una unidad morfológica de todo ser vivo: es decir, que en los seres vivos todo está formado por células o por sus productos de secreción.
-
Este primer postulado sería completado por Rudolf Virchow con la afirmación Omnis cellula ex cellula, la cual indica que toda célula deriva de una célula precedente (biogénesis). En otras palabras, este postulado constituye la refutación de la teoría de generación espontánea o ex novo, que hipotetizaba la posibilidad de que se generara vida a partir de elementos inanimados.
-
Un tercer postulado de la teoría celular indica que las funciones vitales de los organismos ocurren dentro de las células, o en su entorno inmediato, y son controladas por sustancias que ellas secretan. Cada célula es un sistema abierto, que intercambia materia y energía con su medio. En una célula ocurren todas las funciones vitales, de manera que basta una sola de ellas para tener un ser vivo (que será un ser vivo unicelular). Así pues, la célula es la unidad fisiológica de la vida.
-
Finalmente, el cuarto postulado de la teoría celular expresa que cada célula contiene toda la información hereditaria necesaria para el control de su propio ciclo y del desarrollo y el funcionamiento de un organismo de su especie, así como para la transmisión de esa información a la siguiente generación celular.
Características estructurales
- Individualidad: Todas las células están rodeadas de una envoltura (que puede ser una bicapa lipídica desnuda, en células animales; una pared de polisacárido, en hongos y vegetales; una membrana externa y otros elementos que definen una pared compleja, en bacterias Gram negativas; una pared de peptidoglicano, en bacterias Gram positivas; o una pared de variada composición, en arqueas)8 que las separa y comunica con el exterior, que controla los movimientos celulares y que mantiene el potencial de membrana.
- Contienen un medio interno acuoso, el citosol, que forma la mayor parte del volumen celular y en el que están inmersos los orgánulos celulares.
- Poseen material genético en forma de ADN, el material hereditario de los genes y que contiene las instrucciones para el funcionamiento celular, así como ARN, a fin de que el primero se exprese.
- Tienen enzimas y otras proteínas, que sustentan, junto con otras biomoléculas, un metabolismo activo.

La existencia de polímeros como la celulosa en la pared vegetal permite sustentar la estructura celular empleando un armazón externo.
Características funcionales Las células vivas son un sistema bioquímico complejo. Las características que permiten diferenciar las células de los sistemas químicos no vivos son:
- Nutrición. Las células toman sustancias del medio, las transforman de una forma a otra, liberan energía y eliminan productos de desecho, mediante el metabolismo.
- Crecimiento y multiplicación. Las células son capaces de dirigir su propia síntesis. A consecuencia de los procesos nutricionales, una célula crece y se divide, formando dos células, en una célula idéntica a la célula original, mediante la división celular.
- Diferenciación. Muchas células pueden sufrir cambios de forma o función en un proceso llamado diferenciación celular. Cuando una célula se diferencia, se forman algunas sustancias o estructuras que no estaban previamente formadas y otras que lo estaban dejan de formarse. La diferenciación es a menudo parte del ciclo celular en que las células forman estructuras especializadas relacionadas con lareproducción, la dispersión o la supervivencia.
- Señalización. Las células responden a estímulos químicos y físicos tanto del medio externo como de su interior y, en el caso de células móviles, hacia determinados estímulos ambientales o en dirección opuesta mediante un proceso que se denomina quimiotaxis. Además, frecuentemente las células pueden interaccionar o comunicar con otras células, generalmente por medio de señales o mensajeros químicos, como hormonas, neurotransmisores, factores de crecimiento en seres pluricelulares en complicados procesos de comunicación celular y transducción de señales.
- Evolución. A diferencia de las estructuras inanimadas, los organismos unicelulares y pluricelulares evolucionan. Esto significa que hay cambios hereditarios (que ocurren a baja frecuencia en todas las células de modo regular) que pueden influir en la adaptación global de la célula o del organismo superior de modo positivo o negativo. El resultado de la evolución es la selección de aquellos organismos mejor adaptados a vivir en un medio particular.
.
Las enzimas, un tipo de proteínas implicadas en el metabolismo celular

Tipos de células
-
1. Células procariotas
-
2. Células eucariotas
Células procariotas:
Las células procariotas son pequeñas y menos complejas que las eucariotas. Contienen ribosomas pero carecen de sistemas de endomembranas (esto es, orgánulos delimitados por membranas biológicas, como puede ser el núcleo celular). Por ello poseen el material genético en el citosol. Sin embargo, existen excepciones: algunas bacterias fotosintéticas poseen sistemas de membranas internos. También en el Filo Planctomycetes existen organismos como Pirellula que rodean su material genético mediante una membrana intracitoplasmática y Gemmata obscuriglobus que lo rodea con doble membrana. Esta última posee además otros compartimentos internos de membrana, posiblemente conectados con la membrana externa del nucleoide y con la membrana nuclear, que no posee peptidoglucano.
Por lo general podría decirse que los procariotas carecen de citoesqueleto. Sin embargo se ha observado que algunas bacterias, como Bacillus subtilis, poseen proteínas tales como MreB y mbl que actúan de un modo similar a la actina y son importantes en la morfología celular. De gran diversidad, los procariotas sustentan un metabolismo extraordinariamente complejo, en algunos casos exclusivos de ciertos taxa, como algunos grupos de bacterias, lo que incide en su versatilidad ecológica. Los procariotas se clasifican, según Carl Woese, en arqueas y bacterias.
Arqueas

Bacterias

Células eucariotas
La célula eucariota posee compartimentos internos delimitados por membranas. Entre éstos se encuentra el núcleo, delimitado por una doble unidad de membrana, en cuyo interior se encuentra el material genético o ADN que contiene la información necesaria para que la célula pueda llevar a cabo todas las tareas para su supervivencia y reproducción. Entre el núcleo y la membrana plasmática se encuentra el citosol, un gel acuoso que contiene numerosas moléculas que intervienen en funciones estructurales, metabólicas, en la homeostasis, en la señalización. Cabe destacar los ribosomas para la producción de proteínas, el citoesqueleto para la organización interna de la célula y para su movilidad, numerosos enzimas y cofactores para el metabolismo y muchas otras moléculas más. En el citosol también se encuentran los orgánulos, que son compartimentos rodeados por membrana que llevan a cabo funciones como la digestión, respiración, fotosíntesis, metabolismo, transporte intracelular, secreción, producción de energía, almacenamiento, etcétera. Las mitocondrias, los cloroplastos, los peroxisomas, los lisosomas, el retículo endoplasmático, o las vacuolas, entre otros, son orgánulos. El citoplasma es el citosol más los orgánulos que contiene.
Estas se encuentras de manera tradicional dividida en dos tipos células animal y vegetal.
 Esquema de los principales componentes de una célula animal.
Esquema de los principales componentes de una célula animal. 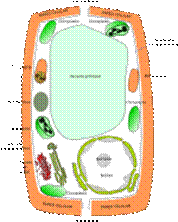 Esquema de los principales componentes de una célula vegetal.
Esquema de los principales componentes de una célula vegetal.

Diagrama de una célula animal, a la izquierda (1. Nucléolo, 2. Núcleo, 3. Ribosoma, 4. Vesícula, 5. Retículo endoplasmático rugoso, 6. Aparato de Golgi, 7. Citoesqueleto (microtúbulos), 8. Retículo endoplasmático liso, 9. Mitocondria, 10. Vacuola, 11. Citoplasma, 12. Lisosoma. 13.Centríolos.).

Estructura de la célula
Membrana Celular
No solo es el límite entre la célula o su entorno sino que también media al contacto y al intercambio entre el citoplasma de una célula y su medio circundante a través de estructuras de transporte. Las membranas celulares son estructuras muy importantes para las células, ya que funcionan como una barrera entre los componentes de la célula y el medio ambiente exterior. La membrana celular no sólo es responsable de crear un muro entre el interior y exterior de la célula, sino que también debe actuar como un umbral a través del cual las moléculas de selección pueden entrar y salir de la celuda cuando sea necesario. La membrana celular es lo que define a la célula y mantiene sus componentes por separado a partir de células u organismos fuera. La membrana celular está compuesta por una doble capa de células de grasa llamado una bicapa lipídica en la que las proteínas de membrana están integradas. La estructura de la bicapa lipídica impide el libre paso de la mayoría de las moléculas dentro y fuera de la célula.
- a. Bicapa lipídica: La estructura de la bicapa lipídica, explica su función como barrera. Los lípidos son grasas, como el petróleo, que son insolubles en agua. Hay dos importantes regiones de un lípido que proporcionan la estructura de la bicapa lipídica. Cada molécula lipídica contiene una región hidrofílica, también llamada región de la cabeza polar, e hidrofóbicos, o región de la cola no polar. La región hidrofílica se siente atraído por las condiciones del agua acuosa, mientras que la región hidrofóbica es repelido por estas condiciones. Dado que una molécula lipídica contiene regiones que son polares y no polares, se llaman moléculas anfipáticas. La clase más abundante de moléculas de lípidos en las membranas celulares es el fosfolípido. Grupo de la molécula de fosfolípidos en la cabeza polar contiene un grupo fosfato. También tiene dos grupos no polares de ácido graso de cadena como la cola. La cola de ácido graso se compone de una cadena de carbonos e hidrógenos. Cuenta con una torcedura en una de las cadenas debido a su estructura de doble enlace. Otra propiedad importante de la bicapa lipídica es su fluidez, la cual permite que estas estructuras de movilidad dentro de la bicapa lipídica. Esta fluidez es de importancia biológica, que influyen en el transporte de membrana. La fluidez depende tanto de la estructura específica de las cadenas de ácidos grasos y la temperatura (aumenta la fluidez a bajas temperaturas). Estructuralmente, la bicapa lipídica es asimétrica: la composición de lípidos y proteínas en cada una de las dos capas es diferente.

- b. Proteínas de la membrana: Además de la bicapa lipídica, la membrana de la célula también contiene una serie de proteínas. Ya hemos mencionado la presencia de ciertas proteínas en la membrana celular. Mientras que la bicapa lipídica aporta la estructura de la membrana celular, las proteínas de membrana permiten muchas de las interacciones que ocurren entre las células. Como ya comentamos en el apartado anterior , las proteínas de membrana son libres de moverse dentro de la bicapa lipídica, como resultado de su fluidez. Aunque esto es cierto para la mayoría de las proteínas, también puede limitarse a ciertas áreas de la bicapa con las enzimas. Las proteínas de membrana realizar diversas funciones, y esta diversidad se refleja en los tipos muy diferentes de las proteínas asociadas con la bicapa lipídica.
Clasificaciones de proteínas de membrana Las proteínas se suele dividir en las clasificaciones más pequeña de las proteínas integrales, proteínas periféricas, proteínas y lípidos de ruedas.
- Las proteínas integrales: Proteínas integrales están incrustadas en la bicapa lipídica. No se puede quitar fácilmente de la membrana celular sin el uso de detergentes que destruyen la bicapa lipídica. Proteínas integrales de flotar libremente y no dentro de la bicapa, al igual que los océanos en el mar. Además, las proteínas integrales son generalmente proteínas transmembrana, que se extiende a través de la bicapa lipídica de manera que un contacto final en el interior de la célula y los otros detalles del exterior. El tramo de la proteína integral, en el interior hidrofóbico de la bicapa es hidrofóbica, compuesto por no-polar aminoácidos. Al igual que la bicapa lipídica, los extremos expuestos de la proteína integral son hidrófilas. Cuando una proteína atraviesa la bicapa lipídica que adopta una configuración de hélice alfa. Proteínas transmembrana puede cruzar la bicapa lipídica o varias veces. El primero se conoce como una sola pasada las proteínas y la tarde como de múltiples pasadas las proteínas. Como resultado de su estructura, las proteínas transmembrana son la única clase de proteínas que pueden realizar funciones tanto dentro como fuera de la célula.
- Las proteínas periféricas: Las proteínas periféricas se unen a la parte exterior de la bicapa lipídica. Son fácilmente separables de la bicapa lipídica, pueden ser removidos sin dañar la bicapa de ninguna manera. Las proteínas periféricas son menos móviles dentro de la bicapa lipídica.
- Lípidos, proteínas Límite: Lípidos, las proteínas se encuentran obligados por completo dentro de los límites de la bicapa lipídica.

- c. Cadena de oligosacáridos: muchas proteínas de la membrana poseen cadenas de oligosacaridos que sobresalen en el espacio extracelular. Si la porción de carbohidrato de la molécula es menos que la porción proteica la molécula se designa glucoproteína; si la porción de carbohidrato es mayor que la proteína de la molécula se designa proteoglucano. Los oligosacáridos son moléculas constituidas por la unión de dos a nueve monosacáridos cíclicos, mediante enlaces de tipo glucosídico. El enlace glucosídico es un enlace covalente que se establece entre grupos alcohol de dos monosacáridos, con desprendimiento de una molécula de agua.

-
d. Diferenciaciones de la superficie celular: Es muy frecuente que la superficie de las células epiteliales, este modificada para desempeñar funciones especializadas. Estas modificaciones obedecen a 2 razones fundamentales:1.- La principal adaptación es la derivada de la necesidad de aumentar la superficie, que en diferentes tipos celulares se traduce en la presencia de microvellosidades, pliegues basales y placas de membrana.2.- La necesidad de mover sustancias por encima de la superficie se consigue gracias a la existencia de proyecciones celulares móviles, denominadas cilios.
-
Microvellosidades: Son proyecciones digitiformes de la superficie celular, aunque en la mayoría de los epitelios se pueden observar pequeñas vellosidades, estando muy desarrolladas en células con funciones o especializaciones absorbentes del intestino delgado. Las formas se mantienen gracias a la existencia de un haz central de filamentos de actina que recorre cada vellosidad y que además están fijados a la corteza de actina. En las células epiteliales del intestino delgado, este haz central, también se encuentra fijado a la red de actina de las uniones adherentes que se establecen entre las células adyacentes. La membrana celular que recubre las microvellosidades contiene glicoproteinas y enzimas específicas de la superficie celular, implicadas en los procesos de absorción. Dicha membrana se observa al M.E como una cubierta borrosa.
-
Los estereocilios: son variaciones de las microvellosidades, pero mucho más largas, y a pesar de su nombre no tienen nada que ver con los cilios. Se encuentran en células epiteliales que revisten los epidídimos y son los sensores de las células ciliadas cocleares.
-
pliegues basales: Son invaginaciones profundas de la superficie basal de las células. Son especialmente prominentes en células implicadas en el transporte de fluidos o iones y con gran frecuencia están asociados con altas concentraciones de mitocondrias, que son las encargadas de proporcionar energía para dicho transporte. La presencia de pliegues basales y mitocondrias confiere un aspecto estriado al citoplasma basal dando lugar al termino de clas epiteliales estriadas.
-
placas basales: Son zonas rígidas de la membrana apical que sólo se observan en el tracto urinario. Pueden quedar replegadas en el interior de la célula cuando la vejiga urinaria está vacía y desplegarse para incrementar la superficie luminal de la célula, cuando esta está llena.
-
Cilios: Son proyecciones en forma de pelos de 0.25 micras de diámetro que se observan en la superficie de ciertas células especializadas y cuya función es la de impulsar fluidos sobre la superficie celular o la de conferir movilidad a la célula. Cada cilio es una extensión sumamente especializada del citoesqueleto y está compuesta por una estructura organizada de microtúbulos (axonema). Estos microtúbulos están asociados a otras proteínas para producir un movimiento de los filamentos que precisa de energía y cuyo resultado es de batir de un lado para otro. Los túbulos forman 9 dobletes externos o periféricos y poseen unos brazos de dineina que dispuestos cada 24 nm, interaccionan con los dobletes adyacentes, para generar el movimiento ondulante del cilio. Otra proteína, la nexina, establece enlaces mas espaciados (86 nm) entre los microtúbulos, estabilizandolos en su posición.De cada uno de los 9 dobletes externos, surgen a intervalos de 29 nm, unas estructuras a modo de radios, que se proyectan hacia el par central de túbulos. Las proyecciones de la vaina central se encuentran cada 14 nm, y se cree que están implicados en la regulación del patrón de batido ciliar.


- e. Mecanismos de endocitosis: La incorporación de sustancias externas por parte de las células animales es esencial para su supervivencia puesto que son heterótrofas. Ya vimos que algunas moléculas salvan la barrera que supone la membrana plasmática mediante difusión o a través de canales, mediante transportadores o por bombas (Transporte a través de membrana). Sin embargo, hay una manera de incorporar grandes cantidades de moléculas al interior de la célula de una sola vez: endocitosis o incorporación de moléculas englobadas en vesículas. De la misma manera que hay un viaje de ida y fusión de vesículas con la membrana plasmática existe un proceso de formación de vesículas en la membrana plasmática, las cuales se fusionan posteriormente con compartimentos internos, principalmente con los endosomas. En esta incorporación masiva, las moléculas extracelulares pueden entrar al interior de la vesícula de forma inespecífica, en solución, o de forma específica unidas a receptores de membrana. El término pinocitosis se refiere a este tipo de endocitosis inespecífica de moléculas disueltas. Agrupados asi: Vesículas recubiertas de clatrina: Es el principal mecanismo por el que se incorporan proteínas integrales y lípidos de la membrana plasmática, así como macromoléculas extracelulares que generalmente no exceden los 156 nm, incluyendo algunos virus. Caveolas. Se describieron hace unos 50 años por P. Palade gracias imágenes de microscopía electrónica. Son unos pequeñas invaginaciones en la membrana plasmática (45-80 nm) presentes en la mayoría de las células eucariotas que posteriormente se transforman en vesículas. Su membrana se caracteriza por poseer una proteína llamada caveolina, además de proteínas periféricas ancladas a glicosilfosfatidil-inositoles, esfingolípidos (esfingomielina y glicoesfingolípidos) y colesterol. La propia existencia de caveolina hace que las células formen caveolas. Hay de 100 a 200 moléculas de caveolina por caveola y existen diferentes tipos en una sola caveola.Macropinocitosis. Es un proceso mediante el cual se incorporan grandes cantidades de fluido extracelular. En la superficie celular se crean evaginaciones a modo de ola cuyo frente cae sobre la membrana plasmática y se fusiona con ella formando una gran vesícula interna o macropinosoma. Fagocitosis. Es un tipo especial de endocitosis que consiste en la incorporación de partículas de gran tamaño como son bacterias, restos celulares o virus. Este mecanismo lo llevan a cabo células especializadas como son los macrófagos, neutrófilos y las células dendríticas. Un ejemplo claro son los macrófagos que fagocitan a los complejos formados por inmunoglobulinas unidas a otras partículas que pueden ser virus o bacterias. También son los encargados de eliminar miles de glóbulos rojos al día. Los protozoos utilizan este mecanismo para alimentarse.



- f. Moléculas de adhesión celular y contactos celulares: Las moléculas de adhesión son receptores funcionales que se expresan en la membrana celular y participan activamente en múltiples fenómenos fisiológicos y patológicos, como son: la organización de las células animales durante el desarrollo embrionario mediante su diferenciación, migración y localización en órganos y tejidos; en los fenómenos de la hemostasia, como la agregación plaquetaria y la formación de trombos; en la reparación tisular y la cicatrización de las heridas; en la diseminación tumoral o metástasis, y desempeñan un papel fundamental en la migración y activación de los leucocitos en la inmunovigilancia, en el desarrollo de la respuesta inflamatoria y de los mecanismos que intervienen en la respuesta inmune celular. La característica fundamental de estos receptores es la capacidad de transducir señales al interior de la célula y modular cascadas de señales inducidas por diferentes factores de crecimiento. El conocimiento de la regulación de la expresión de estas, su estado de activación en la superficie celular, la distribución celular y tisular y sus posibles interacciones, son de crucial importancia en la comprensión de los mecanismos deacción involucrados en el funcionamiento de las células que participan en la defensa inmunológica. Las moléculas de adhesión son receptores celulares funcionales cuya característica principal es la capacidad de transducir señales al interior de las células en su interacción con sus ligandos o contrarreceptores, desencadenando diferentes eventos funcionales celulares como la expresión génica, cambios fenotípicos de inducción y/o sobreexpresión de determinadas moléculas en la membrana celular, y por lo tanto, cambios en el estado de activación de la célula. También estímulos externos como la acción de las citocinas o la estimulación antigénica pueden provocar cambios intracitoplasmáticos que provoquen estos cambios fenotípicos y la activación celular.
Características de algunas moléculas de la superfamilia de las inmunoglobulinas
|
Designación clásica |
Otras designaciones |
Principales ligandos |
Células que las expresan |
|||
|
ICAM-1 |
CD54 |
LFA-1, Mac-1 |
Linfocitos, macrófagos, células endoteliales activadas y epiteliales |
|||
|
ICAM-2 |
CD102 |
LFA-1 |
Células endoteliales no activadas y leucocitos en reposo |
|||
|
ICAM-3 |
CD50 |
LFA-1, a D b 2 |
Células derivadas de la médula ósea incluyendo a las células de Langherans |
|||
|
PECAM-1 |
CD31 |
PECAM-1
|
Monocitos, neutrófilos, plaquetas, subpoblación de células T, células NK y células endoteliales |
|||
|
VCAM-1 |
CD106 |
VLA-4, a 4 b 7 |
Células endoteliales activadas, macrófagos, células dendríticas y estroma medular |
|||
La extravasación del linfocito en el ganglio linfático ocurre selectivamente al nivel de las vénulas poscapilares endoteliales altas (HEV) presentes en las mucosas del tejido linfoide (igualmente ocurre en las placas de Peyer en el intestino). El desarrollo de estas es una consecuencia de la activación de los linfocitos T y la producción de citocinas como el interferón g. La principal característica de esta extravasación, es la mayor adhesividad del linfocito circulante al endotelio alto durante varios segundos, a diferencia del aplanado, al que solo se asocian durante la fracción de un segundo. La baja afinidad de adhesión hace que por la fuerza del flujo sanguíneo, los linfocitos se separen del endotelio, aunque una pequeña parte pueden adherirse firmemente y a través de las células endoteliales penetren al estroma del ganglio linfático. Esta firme adhesión es posible por la interacción de la L-selectina expresada en los linfocitos T vírgenes con la molécula CD34, proteoglicano sobre las células endoteliales, y con la molécula MadCAM-1 expresada en la mucosa intestinal. Si las células T no interaccionan con el antígeno específico en los ganglios linfáticos, estas salen por los vasos linfáticos eferentes, continúan su recirculación linfática y regresan a la sangre a través del conducto torácico. En el intestino, el tiempo de unión de las células T es más prolongado, lo que provoca que el 50 % de estas penetren en la mucosa.
La inflamación es una compleja serie de reacciones homeostáticas que involucra a los mecanismos inmunológicos humorales y celulares para proteger al organismo. Si esta reacción resulta exagerada o crónica, no cumple su función, y ocurren cambios patológicos.
Se caracteriza por una reacción vascular inicial a un estímulo localizado (reconocimiento antigénico) con liberación de mediadores vasoactivos, una reacción celular de reclutamiento de células inflamatorias (leucocitos inmunocompetentes) que depende de la adhesión leucocitaria, y una reacción tisular en la que los leucocitos liberan mediadores inflamatorios, provocando los efectos deseados (eliminación del antígeno) o no (destrucción tisular).



—————